



Laatste bewerking: oktober 2011

Genetische genealogie: van Adam tot Boekelo
NB: door de voortschrijdende inzichten in de (genografische) DNA-wetenschap
zijn hier vermelde gegevens inmiddels ten dele verouderd.
Ook sommige veronderstellingen zijn inmiddels (2020) aan revisie toe.
Na ontvangst van aanvullend Y-dna onderzoek zal deze pagina opnieuw bewerkt worden.
Ik overwoog daarom het onderzoek af te ronden, tot ik de potentiële mogelijkheden ontdekte van de genetische genealogie: DNA-onderzoek als directe bron voor genealogisch onderzoek. Over de meerwaarde van genetische genealogie en meer specifiek voor de geschiedenis en genealogie van de familie Boekelo gaat deze pagina, van Adam tot Boekelo.
Eerdere versies van dit artikel (v.a. juni 2006) zijn met deze herziene versie komen te vervallen.
Enkele jaren geleden heeft National Geographic Channel een film uitgezonden, waarin de geneticus dr. Spencer Wells de kijkers mee nam op een ontdekkingsreis naar de oorsprong van de mensheid. Hij legt in deze film uit hoe het aan de hand van DNA-onderzoek mogelijk is gebleken de migratiepatronen te achterhalen waarlangs onze verre voorouders zich over de wereld verspreid hebben. Deze film, getiteld Journey of Man - A Genetic Odyssey werd enkele jaren geleden meerdere malen uitgezonden door National Geographic Channel. Niet geheel toevallig overigens, want de National Geographic Society nam deel in het DNA-onderzoek van Spencer Wells, het Genographic Project. Aan dit onderzoek heb ik ook mijn DNA-materiaal afgestaan, waarover ik op het vervolg van deze pagina meer zal schrijven. Wie Journey of Man nog niet eerder heeft gezien, kan de hele film in dertien korte afleveringen bekijken via You Tube
Genetisch gezien zijn wij niet meer dan een mix van het DNA van onze voorouders. Daardoor lijken wij op onze ouders, broers en zussen. Net als in bovenstaand voorbeeld kan er zelfs sprake zijn van opvallende kenmerken die in de hele familie voorkomen en vaak vele generaties herkenbaar blijven.
Begin 2006 geleden hoorde ik over het Genographic Project van The Waitt Family Foundation i.s.m. National Geographic en IBM. Het gaat om een wereldwijd onderzoek, waarin de migratie van de mensheid over de wereld, vanaf het ontstaan van de mensheid, in kaart zal worden gebracht. Gedurende vijf jaar zal bij groepen mensen over de hele wereld DNA-materiaal worden afgenomen.
Enkele begrippen toegelicht
Hoewel het menselijk DNA voor 99,9% bij alle mensen gelijk is, kan voor elk mens op aarde op basis van die 0,1% verschil toch een individueel DNA-profiel worden gemaakt.
Binnen die 0,1% bevindt zich het DNA-materiaal waarop het Genographic Project zich richt. Het zijn stukjes DNA die nagenoeg onveranderd worden doorgegeven van vader-op-zoon (Y-chromosoom of y-DNA) en van moeder-op-kinderen (Mitochondrisch DNA, mtDNA).
Slechts heel af en toe (in het mtDNA vaker dan in het y-DNA) vinden ook in deze stukjes DNA mutaties plaats. Juist de mutaties in deze specifieke stukjes DNA maken het mogelijk om de verspreiding van de mens over de wereldbol in kaart te brengen. We spreken van genetische wegwijzers, in jargon: markers.
Alle markers worden aangeduid met een specifieke benaming, zowel bij het y-DNA (bijv. M168, M89), als bij het mtDNA (bijv. L2, M1). Mutaties die nauw aan elkaar verwant zijn worden in één groep ondergebracht. We spreken dan van een haplogroep. Binnen een haplogroep zijn op verschillende niveaus sub-haplogroepen te onderscheiden. Zo kent de haplogroep R de sub-haplogroepen R1a en R1b, maar zijn ook deze sub-haplogroepen weer onder te verdelen, etc. Variaties binnen een haplogroep of sub-haplogroep worden haplotypen genoemd.
Mijn interesse was gewekt. Gesteld dat mijn haplotype (nagenoeg) overeen zou stemmen met dat van personen uit andere families, wat zou dat dan betekenen? Ik nam mij voor om in dat geval de resultaten van mijn genealogische onderzoek in verband te brengen met genealogische onderzoeksresultaten van andere families, of - als dat niet voorhanden zou zijn - daarnaar zelf onderzoek te gaan doen en de onderzoeksresultaten met elkaar te vergelijken. Wellicht is er sprake van een aantoonbare of veronderstelde herkomst uit eenzelfde geografisch gebied. In het gunstigste geval zou dan zelfs een verre familiaire verwantschap aangetoond kunnen worden. De mogelijkheden van de genetische genealogie gaven mijn onderzoek naar de herkomst van mijn familie Boekelo een nieuwe impuls.
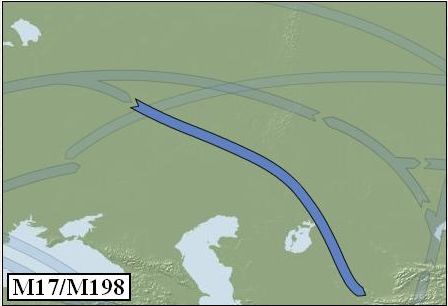
Ik besloot voor USD 126 een DNA-kit te bestellen en deel te nemen aan het project. Blijkbaar was het niet eenvoudig om te bepalen tot welke haplogroep ik me – op basis van het y-DNA onderzoek - mocht rekenen, want er was aanvullend onderzoek nodig. Uiteindelijk ontving ik op 16 juni 2006 de uitslag. Mijn y-DNA haplotype valt binnen de sub-haplogroep R1a1, M17/M198, waarmee sub-haplogroep R1a1 met als kenmerkende markers M17 en M198 wordt aangeduid.

Verschillende betekenissen van R1a1
Recente ontdekkingen hebben er toe geleid dat in de genetisch-genealogische wetenschap verschillende indelingen worden gebruikt om R1* haplogroepen van elkaar te onderscheiden. Dit heeft er toe geleid dat er onduidelijkheid is ontstaan over de gehanteerde benamingen. De meest gebruikte indelingen voor Y-DNA haplogroep R1* zijn die van YCC (2002), Underhill e.a. (2009) en ISOGG (2011).
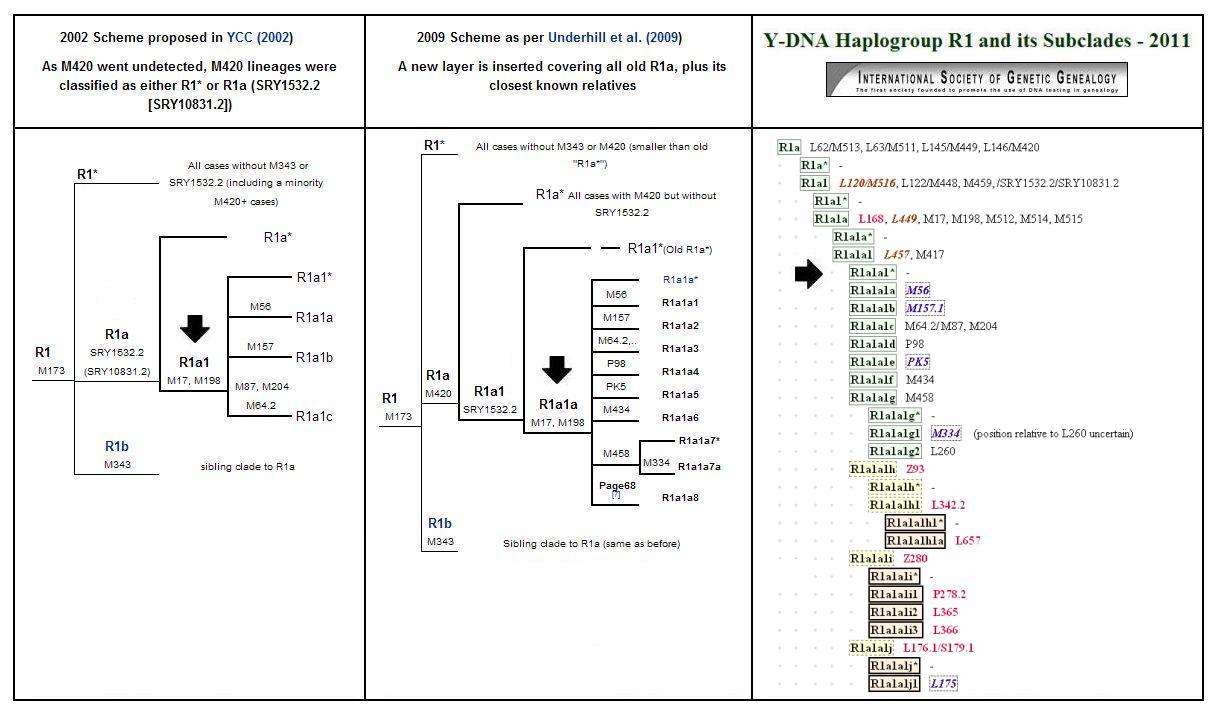
De positie van "mijn" haplogroep wordt in bovenstaande afbeelding aangegeven met een zwarte pijl. In het vervolg van dit artikel gebruik ik de aanduiding R1a1 volgens de indeling van YCC (2002). Bij Underhill (2009) en ISOGG (2011) wordt dezelfde groep respectievelijk aangeduid als R1a1a en R1a1a1*.
In oktober 2011 ontdekte ik op een internet DNA-forum de "haplogroup-predictor" van Vadim Urasin. Met een aan zekerheid grenzende waarschijnlijkheid (99%) geeft deze predictor aan dat mijn haplogroep nader gespecificeerd kan worden als R1a1a1* (M417, -M458, -L365), zoals weergegeven in de indeling van ISOGG (2011).
Vanwege het voorkomen van DYS388=10 (zieonder het kopje Een opmerkelijke STR-waarde op DYS 388 zal ik zeer waarschijnlijk ook positief scoren op L664. Om dat met zekerheid bevestigd te krijgen zal ik nader DNA-onderzoek moeten laten uitvoeren. Dat heb ik nog steeds (oktober 2011) niet gedaan wegens het ontbreken van een volledige 12/12 match, dat verder onderzoek op dit moment weinig zinvol maakt.
Wat mij wel is opgevallen, is dat alle personen die verderop op deze pagina worden genoemd onder het kopje Een vreemd verhaal met 100% zekerheid door Urasins's predictor als R1a1a1* (M417, -M458, -L365) worden benoemd.
Mijn zeldzame en sterk afwijkende waarde DYS439=8 (zie onder het kopje Een zeer zeldzame STR-waarde op DYS439) blijkt overigens niet van invloed te zijn bij de nadere specificatie van Urasin. Het verschil wordt gemaakt door DYS 389(1)=13 in plaats van DYS 389(1)=14.
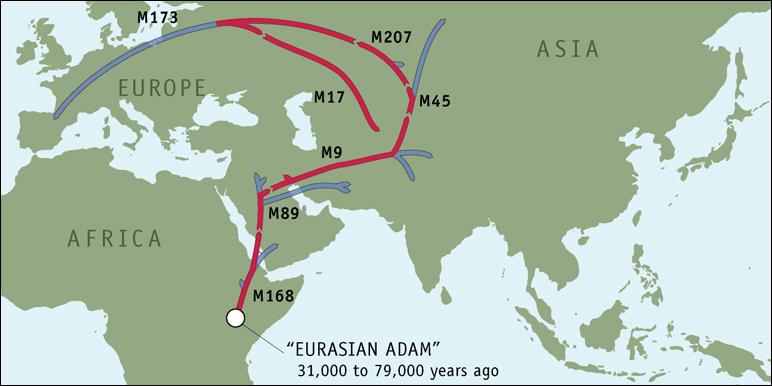
Bij de onderzoeksresultaten die ik ontving was ook onderstaand overzichtskaartje. In grote lijnen geeft het aan hoe de reis is verlopen die mijn verre voorouders hebben gemaakt, van het begin van de mensheid tot aan vandaag de dag. Stap voor stap zal dit verhaal toelichten hoe de reis is verlopen. Een reisverslag, van Adam tot Boekelo.
Fase 1: Out of Africa (M168)
De achtergrondinformatie bij de uitslag van mijn DNA-onderzoek meldt dat de genetische geschiedenis van mijn y-DNA zo'n 50.000 jaar geleden is begonnen met de marker M168.
Kenmerkend voor M168 is dat deze marker is terug te vinden in het DNA van alle niet-Afrikaanse mannen. De mutatie is ergens tussen 31.000 en 79.000 jaar geleden ontstaan, in het gebied van het huidige Sudan en Ethiopië. De mens bij wie deze mutatie voor het eerst is opgetreden noemen we in de genetische wetenschap de Eurasian Adam. Met deze Adam heeft de verspreiding van de mensheid over de wereld – Out of Africa - een aanvang genomen. Geschat wordt dat de wereldbevolking toen uit ongeveer 10.000 mensen heeft bestaan.
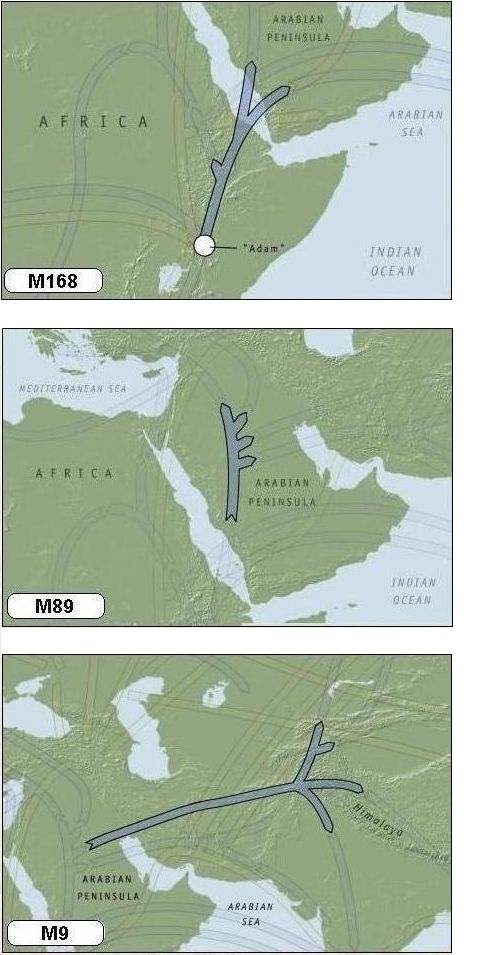
Fase 2: Op het Arabisch schiereiland (M89)
De volgende mutatie die optreedt is in mijn geval marker M89 (Haplogroep F*), die omstreeks 45.000 jaar terug is ontstaan in Noord-Afrika of het Midden-Oosten. De wereld werd toen bevolkt door naar schatting enkele tienduizenden mensen. De marker M89 wordt gevonden in het y-DNA bij 90-95% van alle niet-Afrikaanse mannen.
Mijn verre voorvaderen behoorden tot de tweede golf mensen die het Afrikaanse continent verlieten. Eerder in de geschiedenis was al een andere golf Out of Africa vertrokken, die oostwaarts de kustlijn volgden en uiteindelijk zelfs in Australië terecht gekomen zijn. Mijn voorvaderen trokken echter noordwaarts, over de uitgestrekte grasvlakten die er toen nog in Afrika waren en leefden van de jacht. Al doende trokken zij verder noordwaarts, naar het Arabisch Schiereiland.
Fase 3: Van Arabië naar Azië (M9)
Toen omstreeks 40.000 jaar geleden grote klimaatveranderingen optraden, werd het in het leefgebied van mijn verre voorvaderen kouder en droger. Afrika werd getroffen door droogte en de graslanden veranderden in woestijn. Daardoor konden zij niet meer terug naar Afrika en moesten zij in het Midden-Oosten blijven, of verder trekken.
Velen bleven in het Midden-Oosten, terwijl anderen verder trokken in de richting van het hedendaagse Iran en Centraal Azië. Zij volgden de kuddes wilde dieren, zoals buffalo’s, antilopes, wolharige mammoeten, en andere diersoorten over de halfdroge grasvlakten. In die tijd strekten deze vlakten zich uit van Frankrijk tot Korea en fungeerden als een soort supersnelweg van de oudheid, waarover de mens zich kon verplaatsen.
Omstreeks 40.000 jaar geleden vond ook een nieuwe mutatie in het y-DNA van een van mijn verre voorvaderen plaats. Waarschijnlijk leefde deze man in het huidige Iran of het zuiden van Centraal Azië. Deze mutatie is bekend onder de naam M9 (Haplogroep K).
Nakomelingen van deze voorvader met marker M9 verspreidden zich in 30000 jaar vanuit het huidige Iran en Centraal Azië geleidelijk over grote delen van Eurazië en de Euraziatische steppen.
De genetische wetenschap spreekt in dit verband over de Eurasian Clan. Tot deze groep horen vrijwel alle mensen die van origine afkomstig zijn van het noordelijk halfrond. Tot die groep rekenen we bijna alle Noord-Amerikanen en Oost-Aziaten, evenals de meeste Europeanen en Indiërs. Ondanks het grote verspreidingsgebied van deze groep, werd hun opmars in Azië gestuit door de bergmassieven van zuidelijk Centraal Azië.
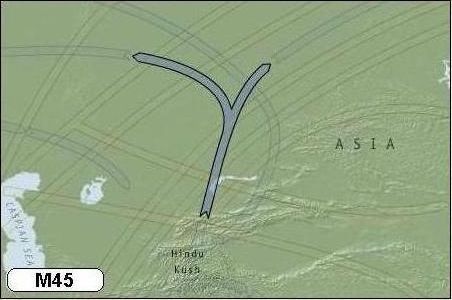
De volgende stap in mijn persoonlijke afstammingsgeschiedenis - langs vaderlijke lijn - brengt mij bij de marker M45 (Haplogroep P). Dit is een van de vier mutaties die is ontstaan uit marker M9.
Marker M45 is 35.000 jaar geleden ontstaan in het y-DNA van een man die geleefd moet hebben in Centraal Azië, ten noorden van het bergachtige Hindu Kush, in het noorden van het huidige Afganistan. De wereldbevolking bestond omstreeks die tijd uit naar schatting 100.000 mensen.
Vanuit Hindu Kush verliep de migratie verder noordwaarts naar de rijke jachtgebieden op de steppes van het huidige Oezbekistan, Kazachstan en Zuid-Siberië. Vindingrijkheid en aanpassingsvermogen waren bittere noodzaak om de IJstijd te kunnen overleven in Siberië, een gebied waar tot dan toe nooit mensachtigen hadden gewoond.
De genetische wetenschap spreekt bij het verschijnen van de marker M45 van het ontstaan van de Central Asian Clan. De voorvader bij wie deze mutatie is ontstaan, is de voorvader geworden van de meeste Europeanen en vrijwel alle oorspronkelijke bewoners van Amerika (in de volksmond: Indianen).
Fase 5: Van Azië richting Europa (M173)
Uit haplogroep P (M45) is vervolgens haplogroep R (M207) ontstaan. Tot deze haplogroep horen de meeste autochtone Europeanen. Wel ontstonden hier uit in de loop der tijd enkele andere haplogroepen.
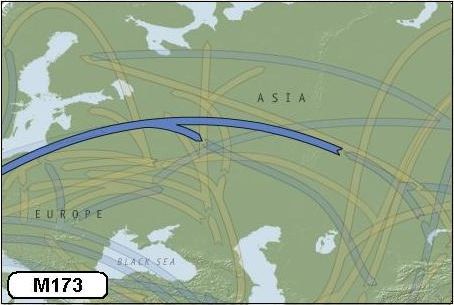
Allereerst was dat de groep met marker M173 (haplogroep R1). De stamvader van deze groep was een man die ongeveer 35.000 jaar geleden in Centraal Azië geleefd moet hebben. Het was de periode dat de Euraziatische steppen zich van het huidige Duitsland - en mogelijk zelf Frankrijk – uitstrekten tot Korea en China. Gunstige klimatologische omstandigheden leidden tot een trek richting Europa. Omstreeks dezelfde tijd trokken deze prehistorische jagers en de nazaten van M173 in een eerste grote golf vanuit Centraal Azië westwaarts Europa binnen.
Vanwege het opdringende landijs werden de nakomelingen van M173 in Europa zo'n 20.000 jaar geleden van lieverlee steeds verder zuidwaarts gedrongen. Daar wisten zij te overleven in de gebieden die wij nu kennen als het Iberisch schiereiland, Italië en de Balkan.
De komst van deze nieuwe Europeanen betekende overigens het einde van de Neanderthalers. Zij vormden een hominide soort, die al in Europa en delen van West-Azië woonden. Betere communicatieve vaardigheden, wapens en de vindingrijkheid van de nieuwkomers hebben er toe geleid dat de Neanderthalers uiteindelijk het onderspit moesten delven.
Toen het klimaat omstreeks 12.000 jaar geleden weer aangenamer werd en het landijs zich weer in noordelijke richting terug trok, trokken ook de prehistorische jagers van M173 weer noordwaarts. Het is dus niet verwonderlijk dat bij veel hedendaagse West-Europeanen de marker M173 in het y-DNA is terug te vinden. We vinden hen in groten getale met name Noord-Frankrijk en de Britse eilanden.
Fase 6: ... en weer terug (M17/M198)
Zo'n 10.000 tot 15.000 jaar geleden ontstonden uit haplogroep R1 (M173) in elk geval twee varianten. We onderscheiden daarbij de haplogroepen R1a1 en R1b. Wat Europa betreft kunnen we in het algemeen stellen R1b (M343) met name in West-Europa voorkomt, terwijl R1a1 (M17/M198) juist meer in Oost-Europa en in mindere mate ook in Noordwest-Europa voorkomt.
Verondersteld wordt dat haplogroep R1a1 zijn oorsprong vindt op de Euraziatische steppen, in het gebied van het huidige Oekraine en Zuid-Rusland. Het is dan tegen het einde van de laatste IJstijd. De wereldbevolking wordt rond die tijd geschat op enkele miljoenen mensen. De R1a1-stamvader en zijn nakomelingen waren nomadische steppebewoners. Wetenschappers opperen een verband met de latere Koergan-cultuur. Mensen die we tot deze cultuur rekenen, zouden als eersten het paard hebben bereden. Daaruit zou dan het grote verspreidingsgebied van R1a1 - van India tot IJsland - verklaard kunnen worden.
Naast genetisch en archeologisch onderzoek is ook linguistisch onderzoek van belang om prehistorische migratiepatronen in kaart te brengen. Sommige taalwetenschappers hangen de theorie aan dat mensen van de Koergan-cultuur zo'n 10.000 tot 5.000 jaar geleden een Proto-Indo-Europese taal spraken. Uit deze taalgroep zijn de latere Indo-Europese talen voortgekomen, zoals de Europese talen maar ook sommige Indiase talen als Bengaals en Hindi.
Tegenwoordig vinden we de grootste concentraties van haplogroep R1a1 in een gebied van Oost-Europa tot aan India, waar 35 procent van de Hindi-sprekende mannelijke bevolking marker M17 draagt.
Hier eindigt het verhaal over de route die mijn voorouders in vaderlijke lijn hebben afgelegd. Naar aanleiding van de uitkomsten van mijn y-DNA onderzoek wilde ik weten wat ik daaruit zou kunnen concluderen over de herkomst van mijn familie langs voorvaderlijke lijn.
De jongste gegevens bevestigen mijn beeld dat ik als West-Europeaan een tamelijk afwijkend y-DNA profiel heb. Waar de meerderheid van de Nederlandse mannen zich mag rekenen tot haplogroepen R1b of I, behoort mijn profiel tot de minderheid R1a1. De percentages R1a1 in Nederland variëren van 3,7% tot 6%.
In Europa komt R1a1 met name voor in de Slavische landen, terwijl R1b vooral in Zuid- en West-Europa veel voorkomt Het zou dus meer voor de hand liggend zijn geweest als ik mij tot deze groep had mogen rekenen. Toch weet ik niet beter of mijn familie woont al eeuwenlang in het Nederlandse en het aangrenzende Duitse taalgebied. Tijd dus voor een nadere beschouwing van de feiten.
Onderstaand overzicht geeft mijn y-DNA profiel weer op basis van 12 onderzochte markers. De markers waarop is getest staan links, de bijbehorende Short Tandem Repeats (STR's) steeds na de dubbele punt. STR's zijn herhalingen van korte DNA-sequenties binnen een bepaalde marker. Mijn profiel ziet er als volgt uit:

Helaas is er tot op dit moment nog niemand in enig publiek toegankelijk y-DNA databestand te vinden met een zogeheten 12/12 match. Dat wil zeggen dat er nog niemand in voor komt met exact hetzelfde profiel als mijn haplotype, waarbij op alle 12 geteste DNA-markers dezelfde aantallen STR's voorkomen. Dit betekent dat er in alle geraadpleegde y-DNA databestanden geen personen voorkomen met wie ook maar een vermoeden van familiaire verwantschap verondersteld kan worden.
Wel vond ik de haplotypen van andere personen, waarvan de genetische afstand enkele mutaties verschilt van mijn haplotype. Dat is te veel om nog te kunnen spreken van een familiaire verwantschap. Die kan pas worden verondersteld bij een 12/12 of soms bij een 11/12 match. Alleen een meer uitgebreide y-DNA test kan daarover uitsluitsel geven.
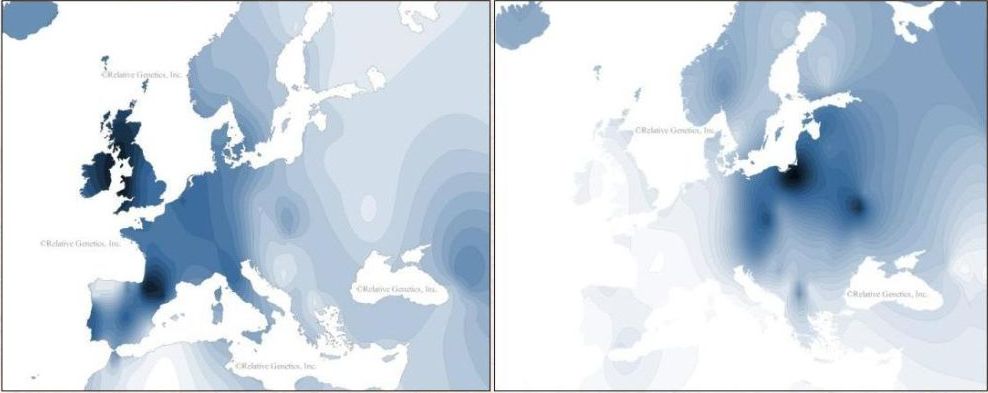
In onderstaand overzicht zijn R1a1 haplotypen weergegeven die het meest verwant zijn aan mijn haplotype. De data zijn afkomstig uit de databestanden van SMGF en Y-SEARCH. Uit deze gegevens lijkt er sprake te zijn van een haplotype dat vermoedelijk ergens in Scandinavië moet zijn ontstaan. Meer uitgebreid genetisch onderzoek is echter nodig om een juist beeld te geven van het voorkomen en de verspreiding van dit haplotype in Europa en met name in Noordwest-Europa; daarover straks meer.
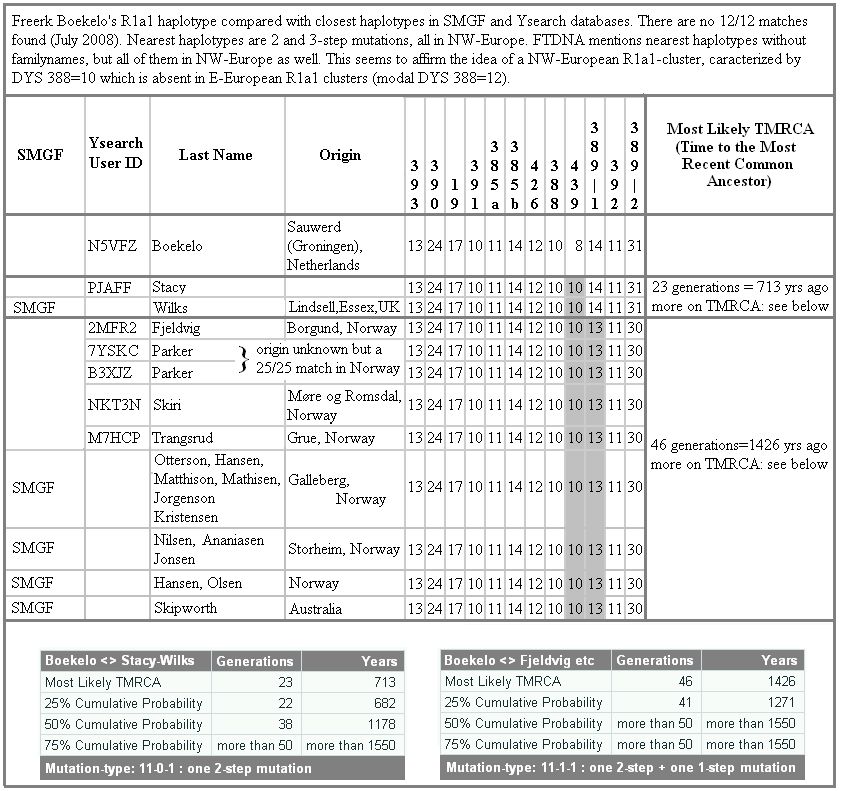
Zoals ik al eerder schreef, verbaasde het mij dat ik behoor tot de sub-haplogroep R1a1, hoofdzakelijk voorkomend in sommige Oosteuropese gebieden en verder oostwaarts. Zoals hiervoor al staat geschreven, is van de mannelijke bevolking van Nederland slechts 3,7%-6% R1a1, terwijl de meeste mannen in ons land zich moeten rekenen tot sub-haplogroep R1b.
Op het internet ben ik vervolgens gaan zoeken naar mogelijke verklaringen voor het voorkomen van R1a1 in Nederland en las daarover het volgende bericht van iemand uit Noorwegen. Hij schreef in oktober 2005 in het Family Tree DNA Forum (vrij vertaald):
 “(…) Om de een of andere reden komt R1a1 maar zelden voor in West-Europa, dus verbaasde me dat een beetje. R1b is namelijk de meest voorkomende haplogroep. Mijn familie woont al generaties lang in Noorwegen (Scandinavië), maar de genetisch kaart laat juist zien dat R1a1 vooral voorkomt in delen van Oost-Europa en in sommige Indiase kasten. (…). Al met al was het nogal verwarrend voor mij, totdat ik las dat in Scandinavië en vooral in West-Noorwegen en op IJsland tot 30% R1a1 voorkomt. Als deze haplogroep opduikt in Ierland en Schotland, dan duidt dit in de regel op Viking-voorvaderen. Het interessante aan deze haplogroep is, dat het voorkomt in verschillende delen van de wereld waar je het niet verwacht.
“(…) Om de een of andere reden komt R1a1 maar zelden voor in West-Europa, dus verbaasde me dat een beetje. R1b is namelijk de meest voorkomende haplogroep. Mijn familie woont al generaties lang in Noorwegen (Scandinavië), maar de genetisch kaart laat juist zien dat R1a1 vooral voorkomt in delen van Oost-Europa en in sommige Indiase kasten. (…). Al met al was het nogal verwarrend voor mij, totdat ik las dat in Scandinavië en vooral in West-Noorwegen en op IJsland tot 30% R1a1 voorkomt. Als deze haplogroep opduikt in Ierland en Schotland, dan duidt dit in de regel op Viking-voorvaderen. Het interessante aan deze haplogroep is, dat het voorkomt in verschillende delen van de wereld waar je het niet verwacht.
Er zijn een hoop theorieën over, maar het is erg interessant. De belangrijkste theorie is dat deze groep mensen de eersten waren die op paarden reden en als eersten een Indo-Europese taal spraken (…). Zij zouden lang geleden andere volken hebben overwonnen. Waarschijnlijk hadden de Vikingen ook voorouders, afkomstig van een deze stammen. Zoals ik al eerder zei: er bestaan vele theorieën, maar niemand weet het zeker.”
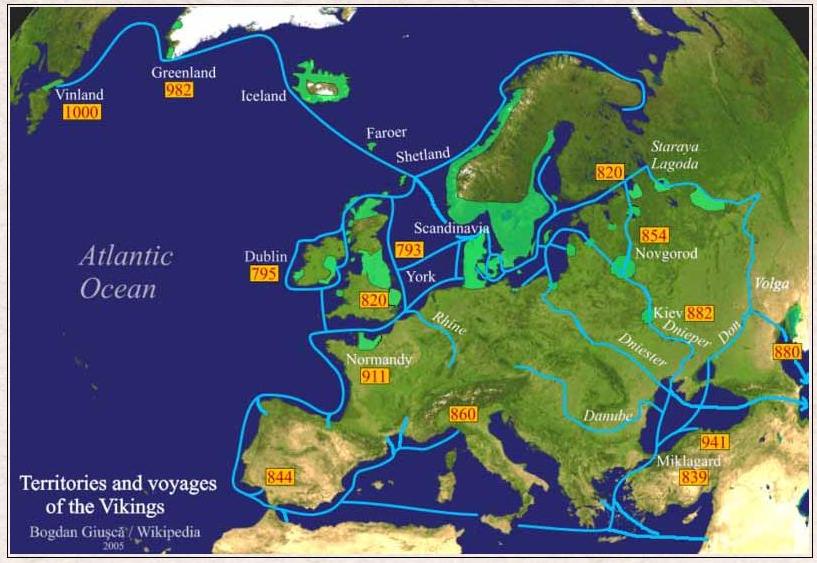
Na lezing van deze posting vroeg ik me af of in mijn geval ook sprake zou kunnen zijn van een Viking-route. De Vikingen zijn immers heel Europa doorgetrokken, ook naar de Lage Landen. Zij zijn vooral berucht geworden vanwege hun rooftochten, maar dreven ook handel in de streken die zij bezochten. Door sommigen wordt deze Vikinglijn zelfs gekoesterd. Zo kunnen inwoners van Engeland, Schotland, Ierland of het eiland Man bij de Oxford Ancestors een certificaat bestellen waarop staat dat zij van de Vikingen afstammen als zij zich mogen rekenen tot haplogroep R1a1. Het voorkomen van deze haplogroep in genoemde gebieden heeft namelijk alles te maken met de activiteiten van de Vikingen in die streken.
Het voorkomen van relatief hoge R1a1 frequenties in sommige delen van Scandinavië (met name in West-Noorwegen) en Oost-Europa waar haplogroep R1a1 vooral in Polen zeer frequent voorkomt, heeft tot speculaties geleid over de herkomst van de oer-Scandinaviërs. Deze speculaties konden tot nu toe nooit wetenschappelijk worden onderbouwd, maar zijn op deze plaats de moeite van het noemen zeker waard.
Zo heeft de bekende - en in wetenschappelijke kringen omstreden - Thor Heyerdahl geprobeerd aan de hand van de IJslandse Sagen aan te tonen dat er ooit sprake is geweest van een volksverhuizing van gebieden in Azië, van achter de Oeral, naar het huidige Noorwegen. De engelstalige website van Wikipedia meldt daarover dat Heyerdahls laatste project wordt beschreven in zijn boek Jakten pĺ Odin ('Op zoek naar Odin'), waarin hij verslag doet van opgravingen in Azov, bij de Zee van Azov in het noordoosten van de Zwarte Zee. Hij zocht daar naar mogelijke overblijfselen van een beschaving die het verhaal van [de middeleeuwse IJslandse schrijver] Snorri Sturluson in zijn Ynglinga Saga zouden bevestigen. Snorri Sturluson beschrijft daarin hoe een stamhoofd met de naam Odin zijn Ćsir-stam voorging in de migratie naar het noorden, door het land van de Saksen, via Fyn in Denemarken naar Zweden, waar de Ćsir-stam zich uiteindelijk zou hebben gevestigd. Snorri verhaalt dat de oorspronkelijke bewoners van het gebied zo onder de indruk waren van Odins kunnen, dat zij hem na zijn dood als een god zijn gaan vereren.
Heyerdahl nam aan dat het verhaal van Snorri op waarheid berustte. Zijn project leverde hem echter een storm van kritiek op van historici, archeologen en taalkundigen in Noorwegen. Zij beschuldigden Heyerdahl van pseudo-wetenschap, van selectief gebruik van bronnen en een groot gebrek aan een wetenschappelijke verantwoorde werkwijze. Heyerdahl concludeert bijvoorbeeld in zijn boek dat bepaalde namen in de Noors mythologie corresponderen met hedendaagse geografische namen in het Zwarte Zee-Gebied, b.v. Azov en Ćsir, Udi en Odin, Tyr en Turkije. Filologen en historici wijzen deze veronderstelling van de hand en spreken van een louter toeval. Zij wijzen er op dat de stad Azov zijn naam pas kreeg omstreeks 1000 jaar nadat Heyerdahls veronderstelde Ćsir-stam in het gebied verbleef.
Blijft de vraag hoe mijn y-DNA profiel, mijn R1a1-haplotype, ooit in het gebied terecht gekomen is waar ik woon en waar al sinds het begin van de 18e eeuw mijn voorouders langs vaderlijke lijn gewoond hebben. Daarover is veel te veronderstellen, maar weinig of niets met zekerheid te zeggen. Het kan inderdaad te maken hebben met de aktiviteiten van de Vikingen tussen 800-1000. Dat zou in elk geval een spectaculair verhaal opleveren voor de familiegeschiedenis of tijdens verjaardagsfeestjes: een Viking-strijder als verre voorvader die ooit in de Lage Landen een inheems meisje bezwangerd heeft .........
Helaas, waarschijnlijk is het allemaal niet zo spectaculair geweest. Ik woon in een gebied waar al sinds mensenheugenis mensen uit alle windstreken hun heil hebben gezocht: soldaten, handelsreizigers, seizoenarbeiders, etcetera. De kans is groot dat ooit één van mijn verre voorvaderen tot deze groep migranten behoorde. Uit den vreemde nam hij zijn voorvaderlijke DNA-profiel mee en gaf het op zijn beurt door aan zijn mannelijke nazaten die niet anders weten dan dat zij van origine uit Nederland komen.
Hoewel over de herkomst van mijn voorvaderlijke familie vooralsnog dus niets met zekerheid gezegd kan worden, heeft vergelijkend y-DNA-onderzoek al wel ěets opgeleverd. Mijn y-DNA bevat namelijk één specifiek kenmerk, waardoor eerder een migratie uit het Scandinavische verondersteld kan worden dan vanuit andere delen van Europa. Daarover gaat het volgende gedeelte.
Een opmerkelijke STR-waarde op DYS 388
Er lijkt sprake te zijn van een Noordwest-Europese variant binnen haplogroep R1a1. Een verband met de Viking-migraties tussen de jaren 800-1000 wordt hierbij al gauw verondersteld, maar migraties uit de pre-Viking periode zijn hierbij evenzeer denkbaar. Er is nog te weinig onderzoek gedaan om hieruit al gefundeerde conclusies te kunnen trekken.
Theorieën over een Noordwest-Europese variant hangen samen met het voorkomen van DYS388=10 binnen haplogroep R1a1. Binnen R1a1 is DYS388=12 het meest voorkomend. Geschat wordt dat ongeveer een-derde tot de helft van alle STR-waarden voor DYS 388=10 binnen haplogroep R1a1 vallen. In een discussie tussen Ken Nordtvedt en Bill Drake op een engelstalig internetforum in januari 2008, werd opgemerkt dat DYS 388=10 op de Britse eilanden met name voorkomt in het zuidwesten van Engeland en Wales, en in mindere mate ook in het zuiden van Engeland, Yorkshire en Ierland. Opvallend is dat DYS 388=10 geheel ontbreekt in het y-DNA van veronderstelde Viking-nakomelingen in het noorden van Groot-Brittannië en op IJsland. Dit duidt er op dat er kennelijk sprake is geweest van verschillende Viking-migratiestromen naar Groot-Brittannië.
Volgens Drake komt DYS 388=10 binnen haplogroep R1a1 in Groot-Brittannië en Scandinavie in ruwweg 5-10% van de gevallen voor. Verder lijkt het er op dat DYS 388=10 in Noorwegen vooral in de kustgebieden voorkomt (Drake verwijst hierbij naar het onderzoek van Dupuy), evenals in België, Nederland, het uiterste noorden van Duitsland, Denemarken en Finland. Met andere woorden, DYS 388=10 komt binnen haplogroep R1a1 met name voor in de kustgebieden van de Noordzee en het Kanaal.
Mogelijk is de mutatie DYS 388=10 ontstaan in het zuiden van Noorwegen of in Denemarken en van daaruit verspreid geraakt langs de kustgebieden van de Noordzee. Het is in elk geval zeker dat de Viking-nakomelingen in het noorden van Groot-Brittannië en op IJsland in genetisch opzicht niet uit een gebied in Noorwegen afkomstig zijn waar de mutatie DYS 388=10 voorkomt.
Een en ander impliceert dat de Viking-invallen in het zuiden van Engeland vooral voor rekening zouden komen van Vikingen uit het zuiden van Scandinavië, zoals bijvoorbeeld de Deense Vikingen. In dat geval zou hetzelfde gelden voor de invallen in het Fries-Groningse gebied.
Aan Bill Drake heb ik de vraag voorgelegd welke conclusie hij verbindt aan het voorkomen van DYS 388=10 in mijn R1a1 profiel. Drake antwoordde in januari 2008 (vertaald): "Van wat ik heb gelezen in openbare databestanden en wetenschappelijke studies, komt DYS 318=10 binnen R1a1 tamelijk veel voor rond Scandinavië en de Noordzee, maar strekt zich niet veel verder uit in de zuidelijke en zuidwestelijke richting van het Europese vasteland. Volgens mij ligt de oorsprong waarschijnlijk ergens rond de Noordzee en mogelijk in Scandinavië. Aangezien deze waarde voorkomt bij ongeveer 5-10 procent van R1a1 in deze regio en naar het zich laat aanzien min of meer gelijkmatig verspreid over gebied voorkomt, zou het kunnen zijn deze mutatie enkele duizenden jaren oud is. Het lijkt hier om een mutatie te gaan met de Scandinavische R1a1 populatie als herkomst.
Ruwweg één-derde van DYS 388=10 binnen R1a1 komt voor in combinatie met DYS 19=15,16 of 17. Er bestaat een subgroep van DYS 388=10 in combinatie met DYS 19=15 en DYS 464=12,14,14,17 waarmee een zekere verwantschap in Engeland bestaat.
Jouw haplotype stemt niet overeen met dit haplotype en het punt waar de genetische splitsing heeft plaatsgevonden ligt waarschijnlijk verder terug in de tijd, mogelijk een paar duizend jaar.
Je Noord-Nederlandse herkomst past goed in het Noordzee-gebied voor wat betreft DYS 388=10 binnen R1a1. Het blijft alleen de vraag wie verantwoordelijk is geweest voor de verspreiding er van. Ligt de herkomst in een ver verleden, dan gaat het zeker om een herkomst die nog vóór de Viking-cultuur gedateerd moet worden. Als DYS 388=10 binnen R1a1 toentertijd al voorkwam in deze gebieden, dan zal het moeilijk, zo niet onmogelijk zijn om vast te stellen of hier sprake is van een vroege datering die terug gaat op een periode vóór de Vikingtijd, dan wel van een latere datering die samenhangt met de komst van de Vikingen vanuit Scandinavië.Meer gegevens over DYS 388=10 zijn eenvoudigweg nog niet bekend, op wat speculaties en onderzoek van enkelen van ons – buitenstaanders – na. Omdat het hier om een minderheidsgroep binnen R1a1 gaat, zijn maar weinig mensen geďnteresseerd in deze groep. We zullen daarom zelf onderzoek moeten doen naar de herkomst van DYS 388=10 binnen R1a1. Het feit dat het hier een kleine groep binnen een minderheids-haplogroep betreft, houdt in dat het nog de nodige tijd zal duren voordat er DNA-testen beschikbaar komen die voor ons doel aangewend kunnen worden."
Lennart Svensson uit Canada heeft zich de afgelopen jaren verdiept in literatuur en databases over het voorkomen van DYS388=10 binnen haplogroep R1a1 in verschillende Noord- en West-Europese landen. Hij concludeert daaruit dat DYS 388=10 met name voorkomt in Noorwegen, Zweden, Denemarken, Sleeswijk-Holstein (Noord-Duitsland), Friesland, Estland en de Britse eilanden (mei 2008)
Paul Gwozdz, deelnemer aan The Polish Project van FTDNA schreef mij (mei 2008) dat van de 422 deelnemers in het project slechts twee personen een STR-waarde van DYS388=10 hadden, maar op andere STR-waarden erg afweken van mijn profiel. Hij concludeert daar uit dat het erg onwaarschijnlijk is dat mijn haplotype tot de Poolse haplotypen binnen R1a1 behoort.
Op het internetforum van DNA-forums.org is Paul Johnsen uit Noorwegen nog stelliger. Hij schreef dat ik vanwege mijn STR-waarde op DYS388=10 met zekerheid ingedeeld moet worden binnen het Noordwest-Europese cluster van sub-haplogroep R1a1. De conclusie van Paul Johnsen lijkt bevestigd te worden door de gegevens in de database van Ysearch (mei 2008): alle scores op DYS388=10 in combinatie met DYS 439=10 (waarover straks meer) in R1a1 vallen binnen het Noordwest-Europese grondgebied:
Op basis van de thans (mei 2010) beschikbare gegevens kan voor DYS 388=10 binnen R1a1 dus gemotiveerd worden verondersteld dat deze STR-mutatie ergens moet hebben plaatsgevonden in het kustgebied van de Noordzee, dan wel in het zuidelijke deel van Scandinavië.
Een zeer zeldzame STR-waarde op DYS439
Een andere opvallende score in mijn profiel is de score op DYS 439, waar een STR-waarde van 8 werd geconstateerd. De meest voorkomende waarde binnen R1a1 op DYS 439=10. De varianten DYS 439=8 en DYS 439=9 komen daarentegen maar zeer sporadisch voor.
Peter Gwozdz en Paul Johnsen wezen mij er op dat er mogelijk sprake is geweest van een multi-step mutation, waarbij in één keer meerdere repeterende stukjes DNA-materiaal zijn gemuteerd. In mijn geval zou dat dus kunnen betekenen dat er een eenmalige mutatie heeft plaatsgevonden van DYS 439=10 naar DYS 439=8. Alleen y-DNA onderzoek van mannelijke familieleden en verre verwanten zou uitsluitsel kunnen geven of dit fenomeen zich inderdaad heeft voorgedaan.
Het verloop van mijn mutaties op DYS 439 kan echter ook anders zijn verlopen. De Y-Chromosome Haplotype Reference Database (YHRD) - release 23 - vermeldt dat de STR-waarde DYS 439=10 bij 3902 haplotypen voor komt. Daarentegen zijn er slechts 11 haplotypen bekend met een STR-waarde van DYS 439=8, terwijl bij 61 haplotypen een STR-waarde is geconstateerd van DYS 439=9.
Op basis van deze frequenties op DYS 439 binnen R1a1, acht ik de mogelijkheid dat DYS 439=8 achtereenvolgens is gemuteerd uit DYS 439=10 en DYS 439=9 ook waarschijnlijk. In dat geval zou DYS 439=8 dus een relatief jonge mutatie zijn.
De vraag is of beide veronderstellingen ook in wetenschappelijk opzicht te verantwoorden zijn. Het antwoord op die vraag moet ik vooralsnog schuldig blijven. Wel vond ik op het internet nog een overzicht van mutatiesnelheden van o.a. de verschillende y-DNA loci waarop ik getest ben. Daaruit blijkt dat mutaties op DYS 439 relatief gezien vaker voorkomen dan op de andere geteste loci (zie afbeelding hierna).

Nieuwe inzichten op ontstaan en verspreiding van haplogroep R1a1 in Europa
Semino e.a. (2000) veronderstellen op basis van hun onderzoek dat haplogroep R1a1 zich aan het einde van de laatste IJstijd (Late Glacial Maximum of LGM) vanuit de Oekraine verspreid heeft over Europa en Azië. Aanvankelijk werd deze hypothese in wetenschappelijke kringen als de meest waarschijnlijke beschouwd. Inmiddels doen andere inzichten de ronde en is de visie van Semino e.a. niet langer leidend in de wetenschappelijke discussies.
Wat betreft de herkomst van haplogroep R1a1 heeft de biochemicus Prof. Anatole A. Klyosov lid van de Russian Academy of DNA Genealogy, een heel andere hypothese ontwikkeld (Klyosov, 2009). Hij stelt dat de gangbare opvatting dat R1a1-M17/M198 omstreeks 15.000 jaar geleden is ontstaan in de huidige Oekraine niet gestaafd wordt door onderzoeksgegevens en daarop gebaseerde berekeningen, maar slechts stoelen op verwijzingen naar andere studies. Klyosov heeft informatie uit openbare DNA-databanken onderzocht en met elkaar vergeleken. Daaruit concludeert hij dat alle Europese mannen die behoren tot haplogroep R1a1, een gemeenschappelijk voorvader hebben die ongeveer 4500-4700 jaar geleden geleefd moet hebben. Een uitzondering maakt hij voor de Balkan (Servie, Kosovo, Macedonië en Bosnië), waar een gemeenschappelijke voorvader ongeveer 11.650 jaar geleden (foutmarge: 1550 jaar) moet hebben geleefd. Een tweede uitzondering betreft de Ierse, Schotse en Zweedse R1a1 populaties, waarbij sprake zou zijn van een veel jongere gemeenschappelijke voorvader in vergelijking met bijvoorbeeld de Russische, Duitse en Poolse R1a1 populaties. Op basis van zijn onderzoek concludeert Klyosov verder dat haplogroep R1a1 zich in Europa zo'n 6000 jaar geleden vanuit de Balkan in verschillende richtingen heeft verspreid.
Ook heeft Klyosov R1a1 haplotypen met de mutatie DYS388=10 geanalyseerd. Hij spreekt van een "subfamily of North-Western European R1a1 haplotypen". Bij ongeveer 20% van de R1a1 haplotypen in Engeland en Ierland komt de mutatie DYS 388=10 voor. De gemeenschappelijke voorvader van deze groep moet omstreeks 3575 jaar geleden geleefd hebben (foutmarge: 450 jaar). Deze DYS 388=10 mutatie komt alleen voor in het noorden en westen van Europa, met name in Engeland, Ierland en Noorwegen en in mindere mate in Zweden, Denemarken, Duitsland en Nederland. Verder naar het oosten en zuiden komt de mutatie DYS388=10 nauwelijks voor. Klyosov heeft in totaal 86 mutaties genoteerd die gebaseerd zijn op een oorspronkelijke R1a1 haplotype met DYS 388=10 (Ysearch-notatie): 13-25-15-10-11-14-12-10-10-13-11-30-15-9-10-11-11-25-14-19-32-12-14-14-17. Volgens zijn berekeningen komt dat overeen met een gemeenschappelijk voorvader die ongeveer 1625 jaar geleden moet hebben geleefd (foutmarge: 240 jaar). Kijkt hij verder terug in de tijd dat komt Klyosov tot 172 mutaties, die terug te voeren zijn op een gemeenschappelijke voorvader die ongeveer 3575 jaar geleden (foutmarge: 450 jaar) geleefd moet hebben: 13-25-16-10-11-14-12-10-10-13-11-30-15-9-10-11-11-24-14-19-32-12-14-15-16. Volgens Klyosov leefde de gemeenschappelijke voorvader van zowel DYS388=12 als DYS388=10 naar alle waarschijnlijkheid op de Balkan, bijna tweeduizend jaar voordat dragers van deze mutatie noord- en westwaarts trokken. Voor DYS 388=12 houdt hij het op 4000 jaar geleden en voor DYS 388=10 op 3600 jaar geleden.
Een ander belangrijk en recent onderzoek is dat van Underhill e.a. (2009). Zij stellen op basis van meer recent onderzoeksmateriaal dat het waarschijnlijker is te veronderstellen dat R1a1 (bij Underhill: R1a1a) zijn oorsprong in Azië heeft. Dat neemt echter niet weg dat deze haplogroep zo'n 11.000 jaar geleden ook al in delen van Europa voorkwam. Veruit de meeste R1a1 haplotypen vallen onder groep R1a1a. Underhill e.a. hebben deze groep R1a1a onderverdeeld in acht subgroepen, van R1a1a1 tot en met R1a1a8, waarvan R1a1a7 (R-M458) de meest voorkomende subgroep in Europa is, met de hoogste frequentie in Polen. Zij veronderstellen dat sub-haplogroep R1a1a7 (R-M458) zich ten tijde van de Steentijd (Neolithicum) met de Touwbekercultuur verspreid heeft over delen van Europa. Andere onderzoekers veronderstellen dat ook het voorkomen van andere R1a1a sub-haplogroepen in Noorwegen in verband gebracht moet worden met de Touwbeker- en Strijdbijlculturen uit Midden- en Oost-Europa. Het begrip Touwbekercultuur is van toepassing op een aantal culturen die verspreid over Europa voorkwamen en leefden op het grensvlak van Steentijd (Neolithicum) en Bronstijd. De naam is ontleend aan de aardewerkvazen die deze culturen fabriceerden en met touwindrukken of visgraatranden versierden. Andere benamingen voor deze cultuur zijn ook wel Strijdbijlcultuur, Strijdhamercultuur, Steenhamervolk, Enkelkamergrafcultuur of Enkelgrafcultuur. In Nederland behoorde de Standvoetbekercultuur tot deze groep.
Het Glazen Lichaam - De geschiedenis in je genen
'Het Glazen Lichaam, een festival dat u inzicht geeft in zowel de
fantastische als de breekbare gevolgen van de modernste
technologieën voor het 'glazen' lichaam in de huidige
informatiemaatschappij. Het Glazen Lichaam is een technologiefestijn
van het Rathenau Instituut en NRC Handelsblad. Het evenement is
gehouden op 2 februari 2008. Het was een avond vol discussies,
presentaties en belevenissen. ' (NRC 3-2-08).

Tot zo ver de voorlopige resultaten en veronderstellingen naar aanleiding van mijn y-DNA onderzoek. Deze pagina zal worden bijgewerkt zodra er meer gegevens bekend worden uit nieuwe onderzoeken. De volledige (engelstalige) tekst van de uitslag van mijn y-DNA-onderzoek is hier te lezen. Voor aanvullingen, opmerkingen en vooral kritische kanttekeningen houd ik mij graag aanbevolen!
Freerk Boekelo
Y-DNA DATABESTANDEN:
YHRD -
YSEARCH -
YBASE -
Oxford Ancestors (OA) -
Relative Genetics (RG) -
Sorenson Molecular Genealogy Foundation (SMGF)
TOOLS:
Y-DNA Haplogroup Tree 2010 -
With Athey's Haplogroup Predictor -
TMRCA calculator -
Y-DNA Comparison Utility
LITERATUUR
Semino et al (2000), The Genetic Legacy of Paleolithic Homo sapiens sapiens in Extant Europeans, Science, Vol 290
Wells et al (2001), The Eurasian Heartland: A continental perspective on Y-chromosome diversity, PNAS, Vol 98
Wells (2002), The Journey of Man: A Genetic Odyssey
Passarino et al (2002), Different genetic components in the Norwegian
population revealed by the analysis of mtDNA and
Y chromosome polymorphisms
Ploski et al (2002), Homogeneity and distinctiveness of Polish paternal lineages
revealed by Y chromosome microsatellite haplotype analysis
Weale et al (2002), Y Chromosome evidence for Anglo-Saxon mass migration
Behar et al (2003), Multiple origins of Ashkenazi Levites: Y chromosome evidence for both
Near Eastern and European ancestries
Capelli et al (2003), A Y chromosome census of the British Isles
Rootsi (2004), Human Y chromosomal variations in European populations
Tambets (2004), The Western and Eastern roots of the Saami—the story of Genetic
“Outliers” told by mitochondrial DNA and Y Chromosomes
Kayser et al (2005), Significant genetic differentiation between Poland and Germany
follows present-day political borders, as revealed by Y-chromosome
analysis
Dupuy et al (2005), Geographical heterogeneity of Y-chromosomal lineages in Norway
McDonald (2005), Y Haplogroups of the World
Pericic et al (2005), High-Resolution Phylogenetic Analysis of Southeastern Europe Traces Major Episodes of Paternal Gene Flow Among Slavic Populations, Society for Molecular Biology and Evolution
McEvoy et al (2006), The scale and nature of Viking settlement in Ireland
from Y-chromosome admixture analysis
Karlsson et al (2006), Y-chromosome diversity in Sweden – A long-time
perspective
Lappalainen et al (2008), Migration waves to the Baltic Sea Region
Wiik et al (2008), Where did European men come from?
Klyosov (2009), DNA Genealogy, Mutation Rates, and Some Historical Evidence Written in the Y-Chromosome, Part II: Walking the Map (In: JOGG, herfst 2009)
Underhill et al. (2009), Separating the post-Glacial coancestry of European and Asian Y chromosomes within haplogroup R1a (In: EJHG, 4 nov. 2009)
WEBSITES
Dienekes' Anthropology Blog
Journal of Genetic Genealogy
American Journal of Human Genetics
European Journal of Human Genetics
National Center for Biotechnology Information - PubMed
The Genographic Project News
Wikipedia
VERWANTE ONDERWERPEN:
Koerganhypothese -
Indo-Europese talen